Adeleorina
Jan Votýpka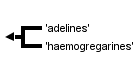

This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxNote: this tree is still under construction. It does not yet contain all known Adeleorina subgroups.
Introduction
Suborder Adeleorina Léger, 1911
Two groups belong to this suborder: (i) “adelines” – monoxenous coccidians of invertebrates, and (ii) “haemogregarines” – heteroxenous coccidians cycling between blood-sucking invertebrates (definitive hosts) and various vertebrates (intermediate hosts). The genus Klossiella (Klossiellidae), a monoxenous coccidium of mammals and reptiles, represents an exception. In veterinary medicine, the most important member representing this group is the genus Hepatozoon, whose member species are mainly pathogenic in domestic mammals such as canids.
Characteristics
Adeleorina are coccidia with the syzygy method of gamete formation, involving the association of often motile gamonts prior to the formation of functional gametes and fertilization. In haemogregarines with heteroxenous species, conjugation of gamonts and subsequent sporogony usually occurs within an invertebrate definitive host which serves as the vector. Similar to all members of the Eucoccidiorida, there is a complex life cycle involving at least one (and often numerous) asexual cycles of merogony followed by gametogony, syngamy, and sporogony. Many species of the group have morphologically distinct types of meronts and merozoites during asexual replication: the first type of meront produces large merozoites which are believed by many to start a further round of merogonic replication; the second type of meront produces smaller merozoites which are said to be progenitors of the gamonts.
The Hemogregarines are important in the history of parasitology. The first species to be seen was apparently Dactylosoma ranarum, by Lankester (1871) in a frog in Europe; he called it Undulina ranarum but later (1882) changed the name to Drepanidium ranarum. So far, there are ~500 named species, almost certainly a great underestimation of the real diversity. Members of the genera Adelina and Adelea infect mostly insects, whereas Klossia is a model coccidium from molluscs.
The haemogregarines (Hepatozoidae, Haemogregarinidae, and Dactylosomatidae) comprise several genera, including pathogens of vertebrates, such as Hepatozoon from carnivores and reptiles, and Haemogregarina from fish and turtles. For all these species, invertebrates play the role of the definitive host with gamogony in their digestive system. Then, basically two modes of transmission occur: (i) the inoculative way (Haemogregarina, Dactylosoma), when the infectious sporozoites enter the vertebrate host during blood-feeding of the vector; (ii) alternatively, the parasite is transmitted by the ingestion of the infected definitive (=invertebrate) host by the appropriate vertebrate host (Hepatozoon, Haemolivia, Karyolyssus). The latter mode of transmission may even involve a paratenic host. Regardless of the mode of transmission, the merogonial division of hemogregarines usually takes place in the parenchymatous organs of vertebrates, followed by the formation of infective gametocytes in the circulating red (in the case of Hepatozoon also white) blood cells. The next definitive host is infected exclusively through blood feeding.
Discussion of Phylogenetic Relationships
Phylogenetic studies indicate that both groups, the monoxenous and heteroxenous coccidians, form a monophyletic clade, characterized also by several morphological and developmental features. Microgamonts produce usually only one to four microgametes, which associate with the macrogamete in syzygy. Other characteristic features of Adeleorina are the absence of endodyogony and the enclosure of sporozoites in a sporocyst.
About This Page
This page is being developed as part of the Tree of Life Web Project Protist Diversity Workshop, co-sponsored by the Canadian Institute for Advanced Research (CIFAR) program in Integrated Microbial Biodiversity and the Tula Foundation.
Jan Votýpka

Department of Parasitology, Charles University
Correspondence regarding this page should be directed to Jan Votýpka at
Page copyright © 2011 Jan Votýpka
 Page: Tree of Life
Adeleorina .
Authored by
Jan Votýpka.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Adeleorina .
Authored by
Jan Votýpka.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 18 May 2011
- Content changed 18 May 2011
Citing this page:
Votýpka, Jan. 2011. Adeleorina . Version 18 May 2011 (under construction). http://tolweb.org/Adeleorina/124811/2011.05.18 in The Tree of Life Web Project, http://tolweb.org/